Какие функции выполняет наружная цитоплазматическая мембрана
Содержание:
Строение
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
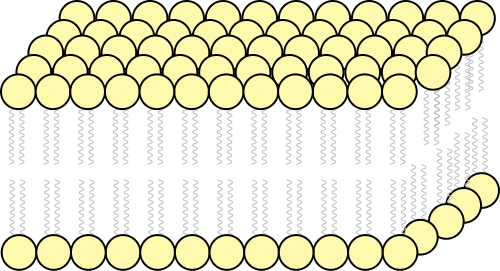
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
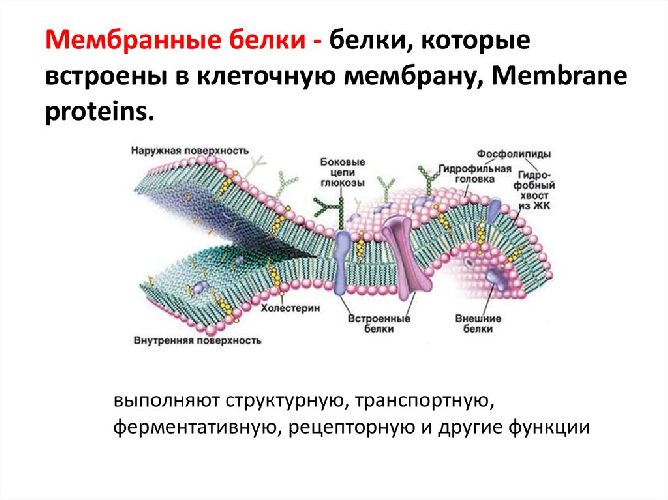
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Структура цитоплазмы
Цитоплазма — внутренняя среда клетки, объединяющая структурные компоненты. Состоит из органелл и цитозоля — «основного вещества» или матрикса (рис. 2). Жидкая фаза цитозоля — коллоидный раствор белковых, минеральных и других веществ. Твердая фаза представлена цитоплазматическим скелетом. Это система трубочек и нитей, постоянно меняющаяся структура, которая создается и разрушается в зависимости от процессов в клетке.
Рис. 2. Строение клетки
Основу цитоскелета составляют:
- Микротрубочки — полые трубки диаметром 20–30 нм, пронизывающие всю цитоплазму.
- Микрофиламенты — нити, образующие сплетения и пучки.
- Промежуточные филаменты — нитевидные образования.
Стенки микротрубочек образованы свернутыми нитями белка тубулина. Сбор белковых молекул для микротрубочек происходит в клеточном центре. Прочные белковые нити образуют опорную основу цитоплазмы. Они противодействуют растяжению и сжатию клетки, поддерживают определенное положение органелл в пространстве. Микротрубочки выполняют опорную и транспортную функцию, так как участвуют в переносе различных веществ.
Микрофиламенты состоят из молекул глобулярного белка актина. Это нити, присутствующие в цитоплазме всех эукариот. Микрофиламенты чаще располагаются вблизи плазматической мембраны, участвуют в изменении ее формы, появлении углублений и выростов
Это особенно важно для пино- и фагоцитоза
Промежуточные филаменты образованы белками, имеют средний диаметр 10 нм (больше диаметра микрофиламентов). Нитевидные структуры тоньше, чем микротрубочки в 2–2,5 раза. Промежуточные филаменты участвуют в создании цитоскелета и движении цитоплазмы.
Клеточные оболочки
Клеточные оболочки
ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной
(синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом
(образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой
(образован целлюлозой).
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны
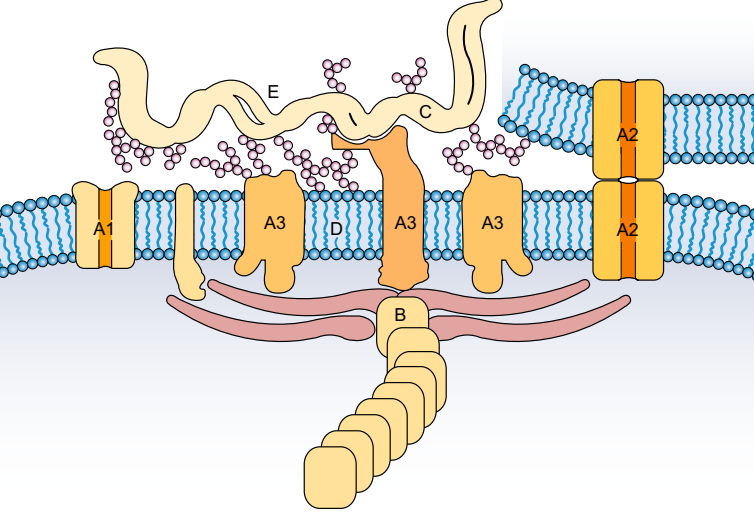
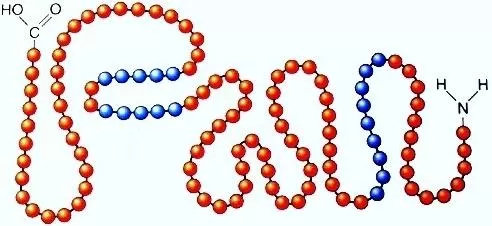
. Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки
(расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки
(погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки
(пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).
А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Избирательная проницаемость
Строение бледной трепонемы
Трехмерная рендер-модель бактерии T. pallidum. Изображены внешняя и цитоплазматическая мембрана (прозрачный желтый), базальные тела (темно-лиловый), аксиальные фибриллы (светло-лиловый), цитоплазматические нити (оранжевый), серповидная «шапочка» возле закругленного конца цитоплазматической мембраны (зеленый), и коническая структура на полюсе (розовый). Пептидогликановый слой не отображен на рендер-модели.
Строение бледной трепонемы (T. pallidum подв. pallidum) было изучено более подробно и изучается в настоящий момент параллельно с развитием иммунологии и электронной микроскопии, начиная с 70-80 годов XX века.
Строение T. pallidum во многом похоже на строение других спирохет.
Проведенные с помощью электронной микроскопии исследования морфологии бледной трепонемы показали, что центральной структурой клетки Т. pallidum является спирально извитой протоплазматический цилиндр.
Протоплазматический цилиндр снаружи окружен цитоплазматической мембраной и плотно прилегающей к нему тонкой клеточной стенкой, основу которой составляет пептидогликан.
Кроме этого, бледная трепонема имеет осевые фибриллы, которые плотно обвиваются вокруг протоплазматического цилиндра. Полагают, что именно они обеспечивают подвижность трепонем, хотя полная функциональность фибрилл не описана в достаточной степени.
Пептидогликан, также известный как муреин, является сложным полимером. Он поддерживает структурную целостность цитоплазматической мембраны и стабилизирует фибриллярный двигательный комплекс. Этот полимер достаточно эластичный, чтобы не мешать сгибательным движениям трепонем.
Бактерия имеет наружную (внешнюю) мембрану. Наружная мембрана охватывает протоплазматический цилиндр и фибриллы.
Аксиальные (осевые) фибриллы располагаются в периплазматическом пространстве, между клеточной стенкой и наружной мембраной. Эти нитчатые структуры тянутся вдоль клетки трепонемы, обвиваясь вокруг её тела в периплазматическом пространстве. Они берут начало от базальных тел, расположенных на обоих концах клетки, и заканчиваются, пройдя середину клеточного цилиндра. Они идут от обоих концов к центру микроорганизма и перекрывают друг друга в центре.
Каждая фибрилла одним концом прикреплена вблизи конца клетки, а другой ее конец свободен. На обоих концах клетки прикреплено одинаковое число фибрилл; в середине или по всей длине клетки фибриллы перекрывают друг друга. В совокупности аксиальные фибриллы называются аксостилем (фибриллярным пучком).
По своим свойствам аксиальные фибриллы напоминают жгутики бактерий. Отличие заключается в том, что аксиальные фибриллы трепонем — внутриклеточные структуры и поэтому называются эндофлагеллами, т.е. внутренними жгутиками.
Т.к. пептидогликановый слой не защищает наружную мембрану, то она легко разрушается при экспериментальных манипуляциях. Фибриллы тоже при этом повреждаются и отстают от тела бактерии, что хорошо видно на многих снимках, полученных в результате электронной микроскопии.
Кроме того, внутри протоплазматического цилиндра также содержатся другие нитевидные структуры, функция которых до сих пор не ясна — цитоплазматические фибриллы, направленные параллельно периплазматическим эндофлагеллам (аксиальным фибриллам).
На концах трепонем наблюдаются структуры конической формы, расположенные в периплазматическом пространстве. По всей видимости, эти уникальные структуры состоят из липопротеинов, упорядоченных в виде спиральной решетки, примыкающей к наружной мембране.
Трепонема, штамм Казань. Электронная микроскопия. K — головная структура. F — фибриллы. F» — цитоплазматические нити.
Срез бледной трепонемы (электронная микроскопия). (ME) — наружная мембрана. (MC) — цитоплазматическая мембрана. (F) — фибриллы. (R) — рибосомы. (N) — вакуоли.
Схема среза бледной трепонемы
Срез концевого сегмента бледной трепонемы на крио-электронной томограмме. Белые треугольники указывают на пептидогликановый слой клеточной стенки, который хорошо видно рядом с концом клетки.
Большинство (50—80%) свежеизолированных штаммов T. pallidum окружены слоем кислых мукополисахаридов, напоминающим капсулу. Однако не все уверены, что это собственный продукт трепонем, а не производное соединительной ткани хозяина. Если это так, то правильнее говорить о псевдокапсуле.
Плазматическая мембрана
Плазмалемма — это перегородка, находящаяся внутри, непосредственно за оболочкой. Она делит клетку на определенные отсеки, которые направлены на компартменты или же органеллы. В них содержатся специализированные условия среды. Клеточная стенка полностью закрывает всю клеточную мембрану. Она выглядит как двойной слой молекул.
Основные сведения
Состав плазмалеммы — это фосфолипиды или же, как их еще называют, сложные липиды. Фосфолипиды имеют несколько частей: хвост и головку. Специалисты называют гидрофобные и гидрофильные части: в зависимости от строения животной или растительной клетки. Участки, которые именуются головкой — обращены внутрь клетки, а хвосты — наружу. Плазмалеммы по структуре являются инвариабельными и очень похожи у различных организмов; чаще всего исключение могут составить археи, у которых перегородки состоят из различных спиртов и глицерина.
Толщина плазмалеммы приблизительно 10 нм.
В малом содержании в состав биологической мембраны входят некоторые виды белков. Например, белки которые пронизывают всю мембрану насквозь, их называют интегральными. Мембраны, которые входят в состав и внешнего, и во внутреннего слоя (слой чаще всего бывает липидным), называются полуинтегральными.
Существуют перегородки, которые находятся на внешней стороне или же снаружи части, вплотную прилегающей к мембране — их называют поверхностными. Некоторые виды белка могут быть своеобразными контактными точками для клеточной мембраны и оболочки. Внутри клетки находится цитоскелет и наружная стенка. Определенные виды интегрального белка могут быть использованы как каналы в ионных транспортных рецепторах (параллельно с нервными окончаниями).
Если использовать электронный микроскоп, то можно получить данные, на основе которых можно построить схему строения всех частей клетки, а также основных составляющих и оболочек. Верхний аппарат будет состоять из трех субсистем:
- комплексное надмембранное включение;
- плазматическая мембрана;
- опорно-сократительный аппарат цитоплазмы, который будет иметь субмембранную часть.
К данному аппарату можно отнести цитоскелет клетки. Цитоплазма с органоидами и ядром называется — ядерный аппарат. Цитоплазматическая или, по-другому, плазматическая клеточная мембрана, находится под клеточной оболочкой.
Слово «мембрана» произошло от латинского слова membrum, которое можно перевести как «кожа» или «оболочка». Термин предложили более 200 лет назад и им чаще называли края клетки, но в период, когда началось использование различного электронного оборудования, установили, что плазматические цитолеммы составляют множество различных элементов оболочки.
Элементы чаще всего структурные, такие как:
- митохондрии;
- лизосомы;
- пластиды;
- перегородки.
Одна из первых гипотез относительно молекулярного состава плазмалеммы была выдвинута в 1940 году научным институтом Великобритании. Уже в 1960 году Уильям Робертс предложил миру гипотезу «Об элементарной мембране». Она предполагала, что все плазмалеммы клетки состоят из определенных частей, по сути, являются сформированными по общему принципу для всех царств организмов.
В начале семидесятых годов XX века было открыто множество данных, на основании которых в 1972 году ученые из Австралии предложили новую мозаично-жидкостную модель строения клеток.
Строение плазматической мембраны
Модель 1972-го года является общепризнанной и по сей день. То есть в современной науке, различные ученые, работающие с оболочкой, опираются на теоретический труд «Строение биологической мембраны жидкостно-мозаичной модели».
Молекулы белков связаны с липидным бислоем и пронизывают всю мембрану полностью — интегральные белки (одно из общепринятых названий — это трансмембранные белки).
Оболочка в составе имеет различные углеводные компоненты, которые будут выглядеть как полисахаридная или сахаридная цепь. Цепь, в свою очередь, будет соединена липидами и белком. Соединенные молекулами белка цепи называются гликопротеинами, а молекулами липидов — гликозидами. Углеводы находятся на внешней стороне мембраны и выполняют функции рецепторов в клетках животного происхождения.
Гликопротеин — представляют собой комплекс надмембранных функций. Его еще называют гликокаликс (от греческих слов глик и каликс, что в переводе означает «сладкий» и «чашка»). Комплекс способствует адгезии клеток.
История исследования
В 1925 году Эверт Гортер и Франсуа Грендель (1897—1969) с помощью осмотического «удара» получили так называемые «тени» эритроцитов — их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем с помощью ацетона выделили из оболочек липиды и определили количество липидов на единицу площади эритроцита — этого количества хватило на сплошной двойной слой. Хотя этот эксперимент привёл исследователей к правильному выводу, ими было допущено несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными плёнками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение — белки. В 1935 году Джеймс Даниэлли и Хью Даусон представили научному сообществу модель «сендвича», которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трёхслойное строение всех клеточных мембран.
Но постепенно накапливались аргументы против «бутербродной модели»:
- накапливались сведения о глобулярности плазматической мембраны;
- оказалось, что структура мембраны при электронной микроскопии зависит от способа её фиксации;
- плазматическая мембрана может различаться по структуре даже в одной клетке, например в головке, шейке и хвосте сперматозоида;
- «бутербродная» модель термодинамически не выгодна — для поддержания такой структуры нужно затрачивать большое количество энергии, и протащить вещество через мембрану очень сложно;
- количество белков, связанных с мембраной электростатически, очень небольшое, в основном белки очень тяжело выделить из мембраны, так как они погружены в неё.
Всё это привело к созданию в 1972 году С. Дж. Сингером и Г. Л. Николсоном жидкостно-мозаичной модели строения мембраны. Согласно этой модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные, полуинтегральные и периферические. Периферические белки действительно находятся на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими взаимодействиями, но никогда не образуют сплошной слой. Доказательствами жидкостности мембраны служат методы FRAP, FLIP и соматическая гибридизация клеток, мозаичности — метод замораживания-скалывания, при котором на сколе мембраны видны бугорки и ямки, так как белки не расщепляются, а целиком отходят в один из слоёв мембраны.
§ 12. Цитоплазматическая мембрана
Строение
Итак, что такое плазма лемма?
Представляет собой тоненькую пленочку разграничивающую прокариоты с внутренней средой. Разглядеть ее можно только в микроскоп. В строение цитоплазматической мембраны входит би слой, который служит основой.
Би слой — это двойная прослойка, состоящая из белков и липидов. Также есть холестерол и гликолипиды, обладают амфипатричностью.
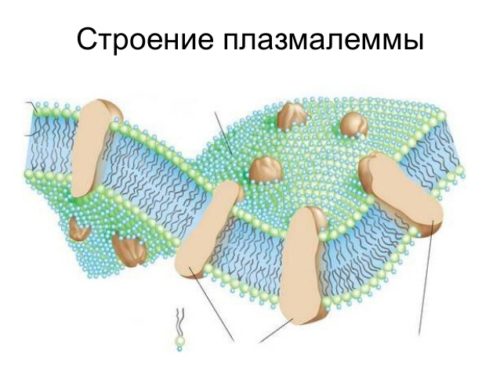 Основой строения цитоплазматической мембраны служит би слой
Основой строения цитоплазматической мембраны служит би слой
Что это значит?
Жировой организм имеет биполярную головку и гидрофильный хвостик. Первая обусловлена боязнью воды, а второй ее поглощением. Группа фосфатов имеет наружное направление от пленки, вторые направлены друг на друга.
Таким образом, происходит формирование биполярного липидного слоя. Липиды обладают высокой активностью, могут перемещаться в своем монослое, редко переходить в другие области.
Полимеры делятся на:
- наружные,
- интегральные,
- пронизывающие плазма лемму.
Первые находятся только на поверхностной части пазухи. Держатся за счет электростатики с биполярными головками липидных элементов. Удерживают питательные ферменты. Интегральные внутри, они встроены в саму структуру оболочки, соединения меняют свое местоположение за счет движения эукариот. Служат своеобразным конвейером, выстроены так, что по ним идут субстраты, продукты реакции. Белковые соединения пронизывающие макрополость имеют свойства образования пор для поступления питательных элементов в организм.
Что такое клеточная мембрана
Если провести аналогию с куриным яйцом (разбив скорлупу, аккуратно отделить от нее тонкую полупрозрачную пленочку), то визуально можно представить, что скорлупа — это плотная клеточная оболочка, а пленка — мембрана. Эта картинка очень наглядно позволяет увидеть, каким образом под клеточной стенкой, состоящей из целлюлозы, располагается плазмалемма. Конечно, это представление будет условным, но, действительно, мембрана в переводе с латинского языка означает кожа. Хотя этот термин достаточно давний, он точно передает сущность мембранной структуры .
Цитолемма (еще одно ее название) животной клетки плотной оболочкой не защищена, однако имеет особый слой, состоящий из белков и жиров, соединенных с сахарами (гликопротеинов и гликолипидов). Называют его гликокаликс, и роль, которую он несет (рецепторная, сигнальная), очень важна для жизнедеятельности.
Строение
Строение структуры уникально, и именно за счет него функции клеточной мембраны выполняются точно и избирательно.
В структуру плазмалеммы входят молекулы:
- фосфолипидов,
- гликолипидов,
- холестерола,
- белков.
Однако не только такой щедрый химический состав делает цитоплазматическую мембрану особой структурой, все свои функции она выполняет благодаря строгой организации молекул.
Строение плазмалеммы физиологически идеально — двойной слой молекул жиров (липидов), полярно организованных, не дают своим выходить за пределы клетки, а чужим — проникать внутрь.
Организация плазмалеммы:
- мембрана состоит из липидов молекулы, которые имеют особое строение,
- каждый липид имеет два конца гидрофильная (любящая воду) головка и гидрофобный (боящийся воды) хвост,
- липиды выстроены таким образом, чтобы головки были снаружи, а гидрофобные хвосты внутри,
- поверхность мембраны гидрофильна (пропускает воду и, соответственно, растворы), а вот внутренняя часть, состоящая из гидрофобных окончаний, воду отталкивает,
- в основном молекулы липидов содержат остатки фосфорной кислоты (это фосфолипиды), некоторые связаны с углеводами (гликолипиды) и холестеролом,
- холестерол придает мембране упругость и жесткость,
- благодаря электростатическим свойствам липиды притягивают молекулы белков, которые также входят в структуру цитолеммы.
Именно белковые молекулы (гранулы) заслуживают отдельного внимания ученых. Из-за своего различного положения и ориентации в полужидкой липидной среде они выполняют самые различные и очень важные функции.
Внутри и на поверхности цитолеммы встречаются следующие виды белков:
- Периферические. Эти молекулы расположены на поверхности и в основном выполняют защитную и стабилизирующую функции. Так, они выстраивают ферменты в конвейерные цепи и не позволяют ферментам просто перемещаться вдоль бислоя.
- Погруженные внутрь (полуинтегральные). Основная их функция — ферментативная, также они могут участвовать в транспорте веществ. Изучена и еще одна интересная роль этих белков — как переносчиков. Они легко соединяются с транспортируемыми молекулами и проводят их внутрь клетки.
- Пронизывающие (интегральные). Они располагаются таким образом, что проходят насквозь, через билипидный слой. Если несколько таких белков сливаются, то образуется канал (пора), через которую могут проходить определенные вещества, связываясь с белковыми молекулами.
Таким образом, все элементы мембранного бислоя несут строго ограниченные своей ролью и строением функции. Благодаря такой организации система работает слаженно и точно.
Отмечено, что плазмалеммы даже внутри одной клетки неоднородны. В них различается не только соотношение химических составных (белков, липидов, углеводов), но и вязкость внутреннего содержимого, ферментативная активность, плотность наружного слоя, толщина.
Месторасположение в клетке
Мембранные структуры буквально пронизывают клеточное содержимое. Они ограничивают все органоиды (за редким исключением, например рибосомы), выстилают их изнутри, являются оболочками ядер.
Самая массивная по содержанию плазмалеммы структура — эндоплазматическая сеть (ЭПР). Если сложить все мембраны, ее составляющие, то получится площадь более половины общей — на все клеточное пространство. По морфологии оболочка ЭПР сходна с внешней ядерной. Они составляют с ней единую систему и обеспечивают активный взаимный перенос элементов.
Комплекс Гольджи — еще один органоид, полностью выполненный из мембранных мешочков (цистерн). Также цитолеммы имеют митохондрии и пластиды.
Плазматическая мембрана — это часть плазмалеммы, находящаяся на границе клеточного содержимого. Она ограничивает протопласт от внешней среды, окружает клетку, защищая его от наружного воздействия.
Основные функции клеточной мембраны
Одним из основных свойств биологических мембран является избирательная проницаемость (полупроницаемость) — одни вещества проходят через них с трудом, другие легко и даже в сторону большей концентрации Так, для большинства клеток концентрация ионов Na внутри значительно ниже, чем в окружающей среде. Для ионов K характерно обратное соотношение: их концентрация внутри клетки выше, чем снаружи. Поэтому ионы Na всегда стремятся проникнуть в клетку, а ионы K — выйти наружу. Выравниванию концентраций этих ионов препятствует присутствие в мембране особой системы, играющей роль насоса, который откачивает ионы Na из клетки и одновременно накачивает ионы K внутрь.
Стремление ионов Na к перемещению снаружи внутрь используется для транспорта сахаров и аминокислот внутрь клетки. При активном удалении ионов Na из клетки создаются условия для поступления глюкозы и аминокислот внутрь ее.
У многих клеток поглощение веществ происходит также путем фагоцитоза и пиноцитоза. При фагоцитозе гибкая наружная мембрана образует небольшое углубление, куда попадает захватываемая частица. Это углубление увеличивается, и, окруженная участком наружной мембраны, частица погружается в цитоплазму клетки. Явление фагоцитоза свойственно амебам и некоторым другим простейшим, а также лейкоцитам (фагоцитам). Аналогично происходит и поглощение клетками жидкостей, содержащих необходимые клетке вещества. Это явление было названо пиноцитозом.
Наружные мембраны различных клеток существенно отличаются как по химическому составу своих белков и липидов, так и по их относительному содержанию. Именно эти особенности определяют разнообразие в физиологической активности мембран различных клеток и их роль, в жизнедеятельности клеток и тканей.
Для многих типов клеток характерно наличие на их поверхности большого количества выступов, складок, микроворсинок. Они способствуют как значительному увеличению площади поверхности клеток и улучшению обмена веществ, так и более прочным связям отдельных клеток друг с другом.
У растительных клеток снаружи клеточной мембраны имеются толстые, хорошо различимые в оптический микроскоп оболочки, состоящие из клетчатки (целлюлозы). Они создают прочную опору растительным тканям (древесина).
Некоторые клетки животного происхождения тоже имеют ряд внешних структур, находящихся поверх клеточной мембраны и имеющих защитный характер. Примером может быть хитин покровных клеток насекомых.